แผนผังง่ายๆ
ของการถ่ายทอดอิเล็กตรอน วัฏจักรเครบส์ส่งอิเล็กตรอน
ผ่านทาง NADH และ FADH2 เข้ามาในระบบถ่ายทอดอิเล็กตรอน
ระบบนี้จะให้พลังงานสำหรับการสร้าง ATP
สัดส่วนใหญ่ของ ATP จากกระบวนการหายใจระดับเซลล์ (cellular
respiration)
มาจากส่วนของออกซิเดทีฟ ฟอสโฟริีเลชัน
:
ท่านทราบไหมว่า เมื่อกลูโคสถูกออกซิไดซ์อย่างสมบูรณ์ให้เป็น
CO2
และ H2O จะได้
ATP จากกระบวนการออกซิเดทีฟ ฟอสโฟริเลชัน
ในสัดส่วนเท่าไรของปริมาณที่จะได้ทั้งหมด
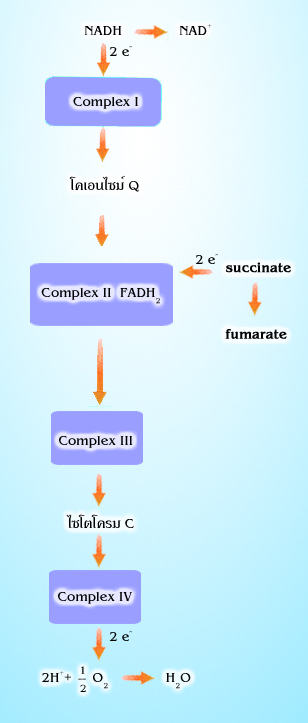
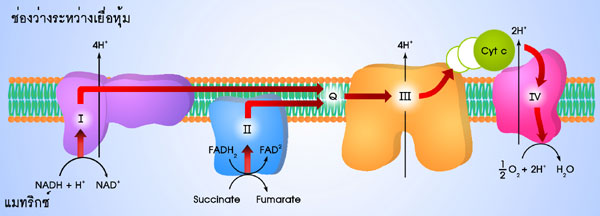
การถ่ายทอดอิเล็กตรอนจะเกิดขึ้นเป็นทอดๆ
ผ่านตัวนำอิเล็กตรอน ซึ่งเป็นกลุ่มของ
โปรตีน (ซึ่งหลายตัวเป็นเอนไซม์) และมีโคเอนไซม์ และโคแฟคเตอร์หลายตัวรวมอยู่ด้วยกัน
กลุ่มโปรตีนเหล่านี้ ได้แก่ complex I, II, III และ IV ลำดับของการถ่ายทอดอิเล็กตรอนได้
แสดงไว้ในรูปที่ 4.2
รูปที่
4.2
แสดงให้เห็นการส่งอิเล็กตรอนเป็นช่วงๆ
ในระบบถ่ายทอดอิเล็กตรอน
(electron transport system ซึ่งบางครั้งเรียกว่า electron
transport
chain หรือ respiratory chain)
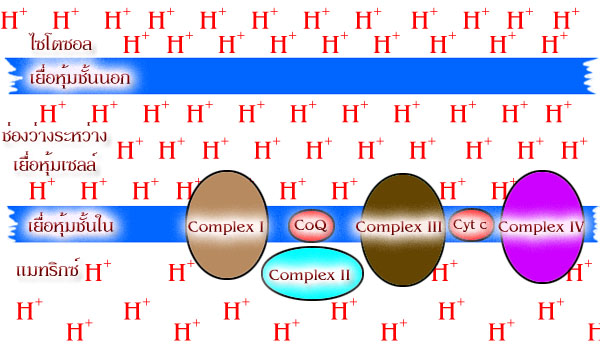
องค์ประกอบของระบบถ่ายทอดอิเล็กตรอน คือ complex I, II, III
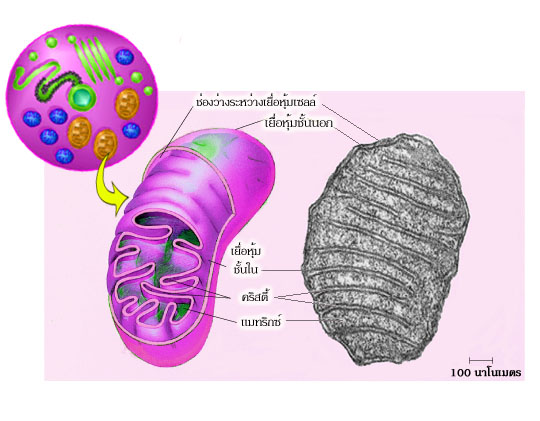
และ IV ฝังตัว
อยู่บนเยื่อหุ้มชั้นในของไมโทคอนเดรีย (ดังแสดงในรูปที่ 4.3) นอกจากกลุ่มโปรตีน
4 กลุ่มนี้
ยังมีโคเอนไซม์ Q และ ไซโตโครม c (cytochrome c ) ซึ่งสามารถเคลื่อนที่ได้เพื่อช่วย
ในการถ่ายทอดอิเล็กตรอนระหว่างกลุ่มโปรตีน ดังกล่าว
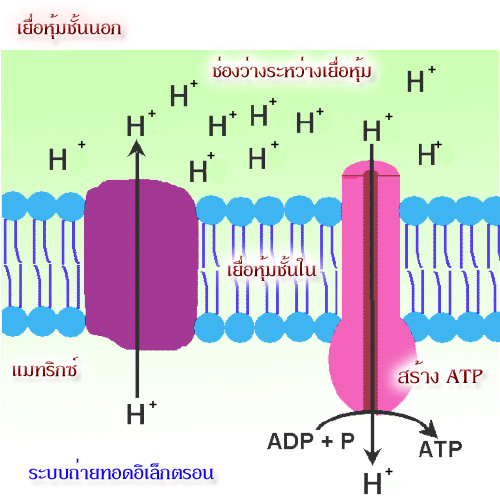
ถึงแม้ว่าจะมีปริมาณโปรตอนที่สะสมอยู่ในทางด้านนอกของเยื่อหุ้มชั้นในจะสูงกว่าทางด้าน
แมทริกซ์มากแต่เยื่อหุ้มนี้จะไม่ยอมให้ H+ ซึมผ่านกลับไปยังด้านแมทริกซ์โดยอิสระได้ตลอดทุกพื้นที่
ของเยื่อหุ้ม แต่ H+จะผ่านไปได้เฉพาะบริเวณจำเพาะที่อยู่บนเยื่อหุ้มชั้นในเท่านั้น
ซึ่งช่อง (channel)
ที่ H+ ผ่านนี้เป็นส่วนหนึ่งของเอนไซม์ ATP synthase
ที่กระจายอยู่ตลอดบนเยื่อหุ้มชั้นใน ขณะที่ H+
ผ่านช่องดังกล่าวเพื่อกลับสู่แมทริกซ์จะเกิดพลังงานที่ใช้สร้าง
ATP ในขณะนั้นเลย
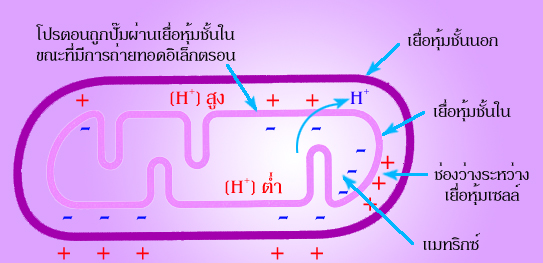
รูปที่
4.6
ภาพแสดงให้เห็นถึงกระบวนการออกซิเดทีฟ
ฟอสโฟริีเลชัน และการสร้าง
ATP ซึ่งเชื่อมโยงกันด้วยการไหลของโปรตอนผ่านเยื่อหุ้มชั้นใน
สรุปได้ว่า
แหล่งพลังงานที่ใช้สร้าง ATP มาจากความแตกต่างของโปรตอนระหว่าง
2 ด้าน
ของเยื่อหุ้มชั้นในของไมโทคอนเดรีย
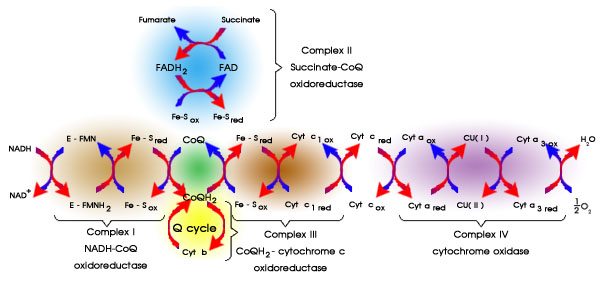
ทุกกลุ่มโปรตีนที่แทรกตัวอยู่ในเยื่อหุ้มชั้นในที่ทำหน้าที่เป็นตัวพาอิเล็กตรอนไม่ว่าจะเป็น
complex I หรือ II หรือ III หรือ IV ประกอบด้วยโปรตีน (เอนไซม์)
หลายตัวที่เกาะอยู่กับหมู่ ที่เรียกว่า prosthetic
group
(โปรดคลิกเพื่อดูรายละเอียด) ที่สามารถถูกออกซิไดซ์และรีดิวซ์ได้ระหว่างการถ่ายทอดอิเล็กตรอน
หมู่เหล่านี้
จำเป็นสำหรับการทำงานของเอนไซม์ เช่น FMN
(flavin mononucleotide) ใน complex I
ซึ่งเป็นโคเอนไซม์ของเอนไซม์ NADH dehydrogenase
นอกจากกลุ่มโปรตีน
I, II, III และ IV ซึ่งแทรกตัวอยู่ในเยื่อหุ้ม ยังมีองค์ประกอบอีก
2 ตัว ที่อยู่บน
เยื่อหุ้มชั้นในแต่สามารถเคลื่อนที่ไปมาได้ คือ โคเอนไซม์
Q (ไม่ใช่โปรตีน เป็นสารโมเลกุลเล็กที่มี
สูตรโครงสร้างที่ทำให้ละลายในไขมันได้ดี (โปรดคลิกเพื่อดูสูตรโครงสร้าง)
ซึ่งจะทำหน้าที่รับอิเล็กตรอน
จาก NADH ผ่านทาง complex I และรับอิเล็กตรอนจาก FADH2
ผ่านทาง complex II
เพื่อส่งให้ complex III ต่อ องค์ประกอบอีกตัวคือ ไซโตโครม
c (cytochrome c) ซึ่งเป็นโปรตีน
ที่มีฮีม (heme) เปรียบได้เหมือนกับฮีโมโกลบินที่เป็นโปรตีนในเม็ดเลือดแดง
แต่ไซโตโครมนี้จะทำหน้าที่
ีรับอิเล็กตรอน ไม่ใช่รับ O2 เหมือนฮีโมโกลบิน โดยที่ไซโตโครม
c จะรับอิเล็กตรอนจาก complex III
เพื่อส่งให้ complex IV ที่มีชื่อว่า ไซโตโครม ออกซิเดส (cytochrome
oxidase) ซึ่งเป็นองค์ประกอบ
ตัวสุดท้ายของระบบถ่ายทอดอิเล็กตรอน ทำหน้าที่เป็นเอนไซม์ที่เร่งปฏิกิริยารีดักชันของ
O2
รูปที่
4.7
แสดงการถ่ายทอดอิเล็กตรอน
จาก NADH ผ่าน complex I และจาก
FADH2 ผ่าน complex II ไปยัง complex III และ
complex IV
โดยมีโคเอนไซม์ Q และ ไซโตโครม c ซึ่งเคลื่อนที่ได้
้เป็นตัวช่วยรับอิเล็กตรอนระหว่าง complex
QH
2 + 2 Cyt ox + 2 H+matrix
Q + 2 Cyt c red + 4 H+cytosol
4)
อิเล็กตรอนถูกส่งจากไซโตโครม
c ไปยัง O2 ผ่าน complex IV
รูปที่
4.12
complex IV รับอิเล็กตรอนจากไซโตโครม
c และถ่ายทอดไปยัง O 2
complex IV คือ ไซโตโครม ออกซิเดส (cytochrome oxidase complex)
หรือ cytochrome a a3
ประกอบด้วย heme A 2 ชนิดคือ a และ a3
ซึ่งเป็นฮีมที่มีทองแดง (Cu) เป็นองค์ประกอบ complex IV
จะรับอิเล็กตรอนมาจากไซโตโครม c และส่งต่อให้ O2 อิเล็กตรอน
4 ตัว จะถูกส่งไปยังโมเลกุลของ O2
เพื่อรีดิวซ์ O2 ให้เป็น H2O ปฏิกิริยานี้เกิดพลังงานอิสระในปริมาณมากสำหรับปั๊มโปรตอนจากด้านแมทริกซ์
ผ่านเยื่อหุ้มชั้นในออกไปด้านที่ติดกับไซโตซอล ซึ่ง proton gradient
ที่เกิดขึ้นนี้ถูกนำมาใช้ในการสร้าง ATP
ปฏิกิริยาที่เกิดขึ้นมีดังแสดงในรูป
4.12 คือ Fe2+ ในไซโตโครม c (ในรูปรีดิวซ์) 2 โมเลกุลส่งอิเล็กตรอนให้
Cu ของไซโตโครม a ซึ่งจะส่งต่อให้ Cu ของไซโตโครม
a3 ซึ่งจะเป็นตัวส่งอิเล็กตรอนต่อให้ O2
รวมปฏิกิริยาที่เกิดขึ้นในขั้นตอนนี้คือ
4
Cyt c red + 8 H+matrix + O
2
4 Cyt c ox + 2 H 2O
+ 4 H+cytosol
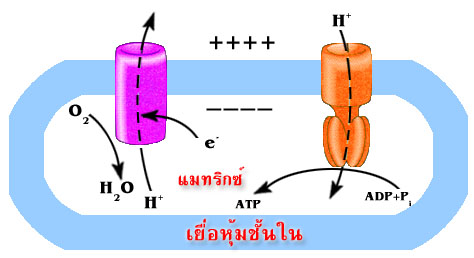
แสดงถึงความสัมพันธ์ระหว่างการปั๊มของโปรตอนผ่านเยื่อหุ้มชั้นใน
จากแมทริกซ์ไปยังช่องว่างด้านนอกที่ทำให้เกิดความแตกต่าง
ของปริมาณโปรตอนใน 2 ด้านของเยื่อหุ้มและการที่โปรตอนไหลกลับ
ผ่านทางช่องจำเพาะของ ATP synthase ทำให้เกิดการสร้าง
ATP
โปรตอนจะไหลกลับเข้ามาในส่วนแมทริกซ์ ผ่านทางโปรตีนที่ฝังอยู่บนเยื่อหุ้มชั้นใน
โปรตีนตัวนี้คือ
ATP synthase ( หรือบางทีเรียกว่า complex V) ATP synthase เป็นเอนไซม์
ประกอบด้วยหน่วยย่อย
หลายหน่วย กลุ่มหนึ่งของหน่วยย่อยทำหน้าที่เป็นช่องให้ H+
ผ่าน อีกกลุ่มหนึ่งทำหน้าที่จับกับ ADP และ
Pi เพื่อสร้าง ATP
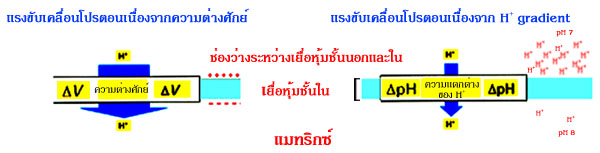
แรงขับเคลื่อนโปรตอนทำให้เกิดการสร้าง ATP
ระบบถ่ายทอดอิเล็กตรอน
และการสร้าง ATP โดย ATP synthase นั้น เป็นคนละระบบแต่เกิดควบคู่กัน
โดยมีแรงขับเคลื่อนโปรตอน (หรือ proton gradient) เป็นตัวเชื่อมโยง
Peter
Mitchell ซึ่งเป็นนักวิทยาศาสตร์รางวัลโนเบลใน คศ.1961 ได้ตั้งสมมุติฐาน
chemiosmotic
เพื่ออธิบายการสร้าง ATP ในไมโทคอนเดรีย โดยกระบวนการออกซิเดทีฟ
ฟอสโฟริเลชัน
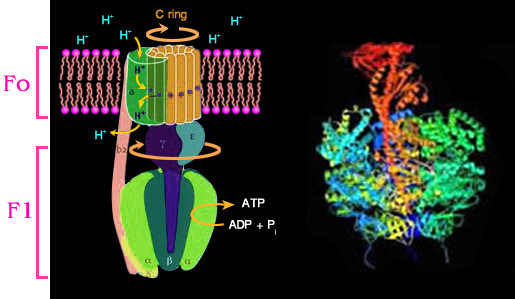
ATP synthase
ประกอบไปด้วย 2 ส่วน มีชื่อเรียกว่า Fo และ F1
โดยที่ส่วน Fo ที่ทำหน้าที่
ลำเลียงโปรตอน (จากด้านนอกของเยื่อหุ้มชั้นในเข้ามาในแมทริกซ์ขณะที่มีการสร้าง
ATP)
และส่วน F1 ทำหน้าที่เร่งปฏิกิริยาสร้าง ATP (catalytic
unit)
หน่วย
Fo แทรกอยู่ในเยื่อหุ้มชั้นใน ประกอบด้วยหน่วยย่อยหลายหน่วยเรียงตัวกันในลักษณะ
ที่ทำให้เกิดเป็นช่อง (channel) สำหรับโปรตอนผ่าน หน่วย Fo
นี้สามารถหมุนได้ ซึ่งจะหมุนต่อเมื่อ
มี H+ ผ่านเยื่อหุ้มชั้นในจากด้านนอกเข้าไปในแมทริกซ์
สำหรับ F1 ซึ่งเป็นส่วนที่อยู่กับที่ เป็นส่วนที่ยื่น
จากเยื่อหุ้มออกมาในแมทริกซ์ หน่วย F1 ต่อกับหน่วย
Fo ด้วยโปรตีนแท่งกลาง
(มี 2 หน่วยย่อย ชื่อ
และ )
F1 ประกอบด้วยหน่วยย่อย 9 หน่วย โดยจะมีหน่วยย่อยที่ชื่อ
จำนวน 3 หน่วย ทำหน้าที่เร่งปฏิกิริยาการสร้าง ATP จาก ADP และ
Pi
รูปที่
4.16
โครงสร้างของเอนไซม์ ATP synthase รูปทางซ้าย: เป็นรูปจำลองอย่างคร่าวๆ
ของเอนไซม์ ATP synthase
ส่วน Fo ฝังอยู่ในเยื่อหุ้มชั้นใน ทำหน้าที่เป็นช่องให้
H+ ผ่าน
ส่วน F 1 ทำหน้าที่เป็นบริเวณเร่งให้ ADP +
Pi ATP
ขณะที่ H+ เคลื่อนที่ผ่านช่องใน F o
ตัว Fo จะหมุนได้ส่งผลให้โปรตีน
แท่งกลางที่เชื่อมต่อระหว่างส่วน Fo และ F1
คือโปรตีน
หมุนได้
และส่งผลต่อไป ทำให้โครงสร้างสามมิติของหน่วย
ย่อยบริเวณเร่งเปลี่ยนไปจนมีการสร้างและปล่อย ATP ออกมาได้ รูปทางขวา: เป็นรูปวาดให้ง่ายขึ้นของภาพสามมิติของ
ATP synthase ซึ่งได้จากการศึกษาทางรังสีเอ็กซ์
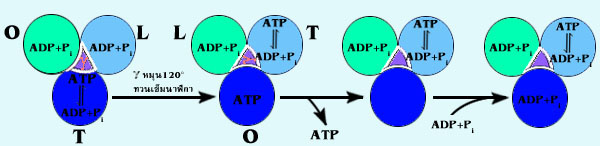
ตามทฤษฎีของ
Paual Boyer ในการสร้าง ATP นั้น 3 หน่วยย่อยที่ชื่อ
จะเป็นตัวจับกับ ADP
และ Pi ได้ทั้ง 3 หน่วย แต่จะเกิดการสร้าง ATP และปล่อยออก
ได้ทีละหน่วย กล่าวคือ หน่วย
ทั้ง 3 หน่วยนี้
จะมีโครงสร้างสามมิติใน 3 แบบ คือ แบบ T (tight) แบบ O (open) และแบบ
L (loose) การเปลี่ยนแปลง
รูปร่างจากแบบหนึ่งไปยังอีกแบบหนึ่ง ทำให้เกิดการเกาะกับ ADP
+ Pi การสร้าง ATP และการปล่อย ATP
แล้วก็กลับไปเริ่มเกาะกับ ADP , Pi ใหม่วนเวียนอยู่เช่นนี้
หน่วยที่มีโครงสามมิติเป็นแบบ
T จะเกาะกับ ATP ได้แน่นมาก ซึ่งทำให้หน่วยนี้สามารถเปลี่ยน ADP,
Pi ให้เป็น ATP แต่ไม่สามารถปล่อย ATP ออกไปได้
หน่วยที่มีโครงสร้างสามมิติแบบ
L สามารถเกาะกับ ADP , Pi ได้ แต่ไม่สามารถเปลี่ยนให้เป็น
ATP ได้
หน่วยที่มีโครงสร้างสามมิติแบบ
O เป็นรูปแบบที่จะปล่อย ATP ออกไปจากตัวและสามารถจับ
กับ ADP + Pi ได้
การเปลี่ยนโครงสร้างสามมิติเกิดขึ้นเนื่องจากการหมุนของโปรตีน
ที่เชื่อมระหว่าง Fo
และ F1
ไป 120 oC ไปในทิศทางทวนเข็มนาฬิกา (เมื่อมองจากด้านบน)
และโปรตีน นี้จะหมุนได้ต่อเมื่อ
มีโปรตอนผ่านส่วน Fo ทั้งนี้โครงสร้างสามมิติของแต่ละหน่วยจะเปลี่ยนไปในทางที่สามารถทำงาน
ต่อเนื่องใน 3 ขั้นตอนของการสร้าง ATP ได้ ในการหมุนของโปรตีน
120 oC ทวนเข็มนาฬิกา หนึ่งครั้ง
จะมี ATP ถูกปล่อยไปหนึ่งตัว ดังแสดงในรูปที่ 4.17
รูปที่
4.17
การทำงานของเอนไซม์ ATPase ตามทฤษฎีของ Paual Boyer
ด้วยกลไก binding – change การหมุนของโปรตีน
(เนื่องจากมี H+ ไหลผ่าน) ทำให้หน่วย
ทั้ง 3 หน่วย เปลี่ยนโครงสร้าง
สามมิติกลับไปมาใน 3 รูปแบบ คือแบบ T ที่มี ATP ที่สร้างใหม่
แต่ปล่อยออกไม่ได้ จะเปลี่ยนเป็นแบบ Oซึ่งปล่อย ATP
ออกไป
และจับ ADP , Pi ตัวใหม่ และเปลี่ยนเป็นแบบ
L เพื่อที่จะจับกับ ADP,
Pi ให้แน่นขึ้นเพื่อจะเริ่มการสร้าง ATP
ในรอบใหม่
จากรูปที่
4.17 หน่วย T (สีน้ำเงิน) เปลี่ยนโครงสร้างสามมิติเป็นแบบ O ทำให้ปล่อย
ATP ที่สร้างขึ้นมา
(ขณะที่เป็น T) ออกไปและมี ADP, Pi ตัวใหม่มาเกาะ ขณะเดียวกันหน่วย
L (สีฟ้า) เปลี่ยนเป็น T
ทำให้เร่งปฏิกิริยา ADP, Pi เป็น ATP แต่ยังไม่สามารถปล่อย
ATP ออกมาได้ ในเวลาเดียวกัน
หน่วย O (สีเขียว) เปลี่ยนเป็น L จับ ADP และ Pi ได้แน่นพอที่จะไม่ปล่อยให้หลุดออกไปเพื่อเตรียม
ที่จะสร้าง ATP เมื่อรูปร่างเปลี่ยนไปเป็นแบบ T อีกครั้งหนึ่ง
โดยสรุป
ATP ถูกสร้างและปล่อยออกมาเมื่อโปรตอนไหลกลับจากช่องว่างระหว่างเยื่อหุ้มชั้นใน
และชั้นนอกมายังแมทริกซ์ผ่านช่อง Fo ของเอนไซม์ ATP
synthase และการสร้าง
ATP เกิดขึ้นที่ส่วน F1
(โปรดคลิกเพื่อดูรายละเอียดของเอนไซม์ ATP
synthase เพิ่มเติม)