
อาร์เอ็นเอชนิดต่างๆ ซึ่งยังอยู่ในสภาพที่ยังไม่พร้อมที่จะถูกนำไปใช้งาน จึงต้องมีการตัดแต่ง
โมเลกุลของอาร์เอ็นเอเหล่านี้เสียก่อนก่อนที่จะนำออกสู่ไซโทพลาซึม โดยการตัดแต่ง
โมเลกุลของอาร์เอ็นเอเหล่านี้จะเกิดขึ้นภายในนิวเคลียส และพิเศษสำหรับ rRNA การตัดแต่ง
โมเลกุลจะเกิดในบริเวณที่จำเพาะที่เรียกว่านิวคลีโอลัส (nucleolus) ขั้นตอนของการตัดแต่ง
นั้นมากบ้างน้อยบ้างขึ้นอยู่กับชนิดของอาร์เอ็นเอ
| การตัดแต่ง tRNA |
Pre-tRNAs
ที่ถูกสังเคราะห์ใน โปรแคริโอต และ ยูแคริโอต เป็นโมเลกุลที่มีลักษณะ
เป็นรูปห่วงเกิดจากการสร้างพันธะระหว่างเบสในสายเดียวกัน กระบวนการตัดแต่ง tRNA
มีขั้นตอนที่สำคัญ 3 ขั้นตอนคือ (1) การตัดนิวคลีโอไทด์บางส่วนทางปลายด้าน 5' โดย
เอ็นไซม์ RNase P ribozyme (อ่านว่า อาร์เอ็นเอส พี ไรโบไซม์) (2) การตัดนิวคลีโอไทด์
บางส่วนทางปลายด้าน 3' โดยเอ็นไซม์ RNase D (อ่านว่า อาร์เอ็นเอส ดี) แล้วเติมเบส
เข้าไปอีก 3 ตัว คือ CCA โดย tRNA ทุกโมเลกุลจะได้รับการตัดแต่งปลายด้าน 3' ให้มีเบส 3 ตัวสุดท้ายเป็น CCA เสมอ และปลายด้านนี้จะเป็นตำแหน่งที่จะให้กรดอะมิโนมาเกาะ เพื่อนำ
ไปใช้ในระหว่างการสังเคราะห์โปรตีน และ (3) เป็นการดัดแปลงเบสบางตัวบนโมเลกุล tRNA
ด้วยวิธีการต่างๆ เช่น การเติมหมู่เมทิล (methylation) หรือการเอาหมู่อะมิโนออก
(deamination) เป็นต้น ตัวอย่างเบสที่ได้รับการดัดแปลงแล้ว ได้แก่ dihydrouridine (D)
ribothymidine (T) pseudouridine ( )
inosine (I) และ 1N-methylguanosine (m1G)
)
inosine (I) และ 1N-methylguanosine (m1G)
จำนวนเบสที่เติมเข้าไปมีประมาณ 10% ของจำนวนเบสทั้งหมด นอกจากนี้ยังมีการตัดเบส
บางส่วนบน tRNA ออกไปด้วย และพิเศษสำหรับยูคาริโอตอีกอย่างหนึ่งคือ ใน tRNA จะมี
อินทรอน (intron) อยู่ด้วยดังนั้นการตัดแต่งโมเลกุลจึงเพิ่มขั้นตอนของการตัดอินทรอนออก
เข้ามาอีกขั้นตอนหนึ่ง
เป็นรูปห่วงเกิดจากการสร้างพันธะระหว่างเบสในสายเดียวกัน กระบวนการตัดแต่ง tRNA
มีขั้นตอนที่สำคัญ 3 ขั้นตอนคือ (1) การตัดนิวคลีโอไทด์บางส่วนทางปลายด้าน 5' โดย
เอ็นไซม์ RNase P ribozyme (อ่านว่า อาร์เอ็นเอส พี ไรโบไซม์) (2) การตัดนิวคลีโอไทด์
บางส่วนทางปลายด้าน 3' โดยเอ็นไซม์ RNase D (อ่านว่า อาร์เอ็นเอส ดี) แล้วเติมเบส
เข้าไปอีก 3 ตัว คือ CCA โดย tRNA ทุกโมเลกุลจะได้รับการตัดแต่งปลายด้าน 3' ให้มีเบส 3 ตัวสุดท้ายเป็น CCA เสมอ และปลายด้านนี้จะเป็นตำแหน่งที่จะให้กรดอะมิโนมาเกาะ เพื่อนำ
ไปใช้ในระหว่างการสังเคราะห์โปรตีน และ (3) เป็นการดัดแปลงเบสบางตัวบนโมเลกุล tRNA
ด้วยวิธีการต่างๆ เช่น การเติมหมู่เมทิล (methylation) หรือการเอาหมู่อะมิโนออก
(deamination) เป็นต้น ตัวอย่างเบสที่ได้รับการดัดแปลงแล้ว ได้แก่ dihydrouridine (D)
ribothymidine (T) pseudouridine (
จำนวนเบสที่เติมเข้าไปมีประมาณ 10% ของจำนวนเบสทั้งหมด นอกจากนี้ยังมีการตัดเบส
บางส่วนบน tRNA ออกไปด้วย และพิเศษสำหรับยูคาริโอตอีกอย่างหนึ่งคือ ใน tRNA จะมี
อินทรอน (intron) อยู่ด้วยดังนั้นการตัดแต่งโมเลกุลจึงเพิ่มขั้นตอนของการตัดอินทรอนออก
เข้ามาอีกขั้นตอนหนึ่ง
 |
: คุณคิดว่าการตัดแต่งโมเลกุลของ tRNA ในขั้นตอนใดมีความสัมพันธ์กับ การแปลรหัสเพื่อการสังเคราะห์โปรตีนมากที่สุด เพราะเหตุใด |
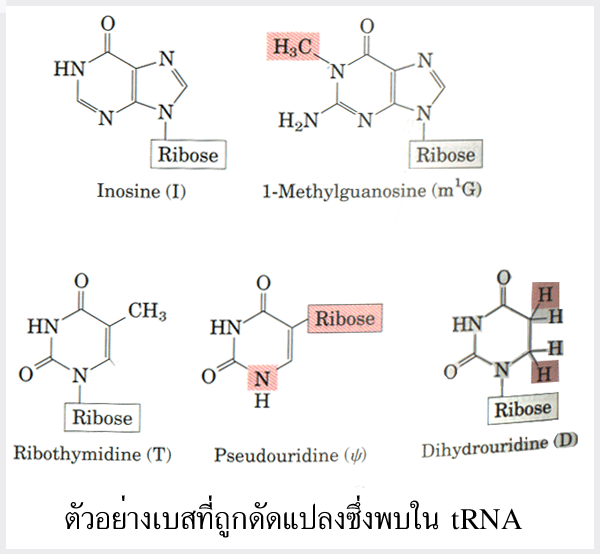
รูปที่ 3.8 ตัวอย่างของเบสที่ถูกดัดแปลงซึ่งพบใน tRNA

รูปที่ 3.9 การตัดแต่ง t-RNA
คล้ายกันคือ การตัดโมเลกุลของ pre-rRNA ให้ได้ขนาดของ rRNA ตามที่ต้องการ โดยใน
โปรแคริโอต โมเลกุลของ pre-rRNA ที่ถูกสังเคราะห์ขึ้นในตอนแรกจะมีขนาด 5.5 kb
ประกอบด้วยลำดับนิวคลีโอไทด์ที่สังเคราะห์มาจากยีนที่จะถูกคัดลอกให้ rRNA 3 ชนิด ได้แก่
16S 23S และ 5S โดยมีขนาด1.5 kb 2.9 kb และ 0.12 kb ตามลำดับ ส่วน pre-rRNA ของ ยูแคริโอต จะมีขนาด 13 kb และสังเคราะห์มาจากยีนที่จะถูกคัดลอกให้ rRNA 3 ชนิด ได้แก่
18S 5.8S และ 28S โดยมีขนาด 1.9 kb 0.16 kb และ 5 kb ตามลำดับ สำหรับ rRNA ของ
ยูแคริโอต อีกชนิดหนึ่ง คือ 5S จะไม่ผ่านกระบวนการตัดเหมือนเช่น rRNA ชนิดอื่น เนื่องจาก
5S-rRNA ถูกคัดลอกมาจากยีนที่แยกอยู่ต่างหาก (ไม่ได้อยู่รวมกับ pre-rRNA ขนาด 13 kb)
rRNAs ของยูแคริโอตที่ถูกตัดให้ได้ขนาดตามที่ต้องการแล้วจะถูกเติมหมู่ methyl ที่เบส
และที่น้ำตาลไรโบสของนิวคลีโอไทด์บางตัว
|
: คุณทราบหรือไม่ว่าตัวพยัญชนะ “S” ที่ปรากฏอยู่ใน rRNA ชนิดต่างๆ เช่น 16S 23S หรือ 5S หมายถึงอะไร |
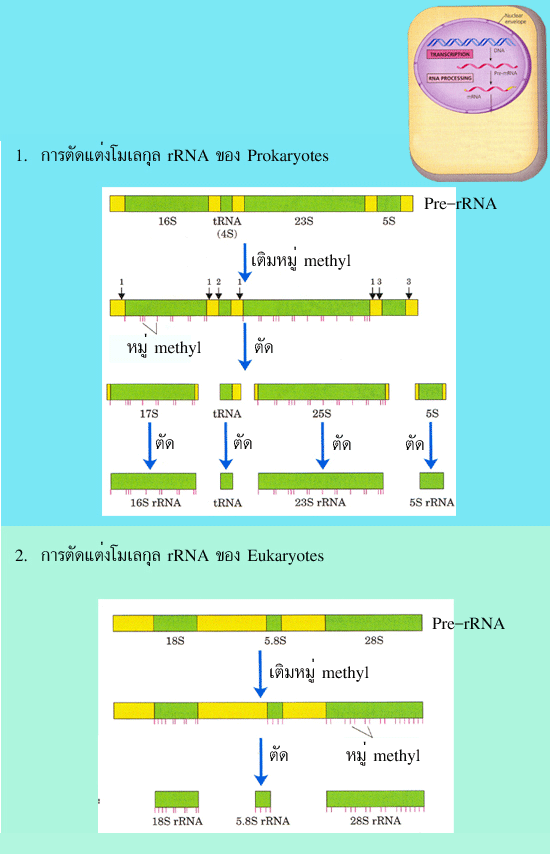
รูปที่ 3.10 เปรียบเทียบการตัดแต่ง rRNA ในโปรแคริโอต และยูแคริโอต
methylguanosine (m7G) ทางปลายด้าน 5' โดยขั้นตอนนี้เกิดขึ้นเมื่อการสังเคราะห์ mRNA
ผ่านไปได้ประมาณ 20-30 นิวคลีโอไทด์ และเกิดโดยอาศัยคุณลักษณะพิเศษของเอ็นไซม์
RNA polymerase II (คุณลักษณะพิเศษดังกล่าวนี้ไม่พบในเอ็นไซม์ RNA polymerase I และ
III ดังนั้นจึงไม่เกิดการเติม m7G ที่ปลายด้าน 5' ของ rRNA และ tRNA) (2) การเติม
นิวคลีโอไทด์ที่มีเบส A (poly-A tail) ประมาณ 50-250 นิวคลีโอไทด์ ทางปลายด้าน 3'
โดยก่อนการเติม poly-A tail ปลายด้าน 3' ของ pre-mRNA จะถูกตัดบางส่วนออกก่อน โดย
ตำแหน่งที่จะถูกตัดจะอยู่ห่างจากลำดับนิวคลีโอไทด์ที่เป็นสัญญาณการตัด (AAUAAA) ออก
ไปทางด้าน 3' ประมาณ 10-30 นิวคลีโอไทด์ และ (3) การตัดอินทรอนซึ่งอยู่ภายในสายของ
mRNA ออกและเชื่อมเอ็กซอน (exon) เข้าด้วยกัน โดยในขั้นตอนที่ 1 มีวัตถุประสงค์เพื่อ
ประโยชน์ในการจดจำ และเกิดการจับระหว่าง mRNA กับไรโบโซม และป้องกัน mRNA
ไม่ให้ถูกทำลายด้วยเอ็นไซม์ ส่วนในขั้นตอนที่ 2 เป็นการป้องกัน mRNA ไม่ให้ถูกทำลาย
ด้วยเอ็นไซม์ และยังเอื้อประโยชน์ต่อการนำ mRNA จากนิวเคลียสออกสู่ไซโทพลาซึมด้วย
และขั้นตอนที่ 3 เป็นการตัดส่วนที่ไม่จำเป็นในการสังเคราะห์โปรตีนออก
โปรแคริโอต ไรโบโซมจะจดจำ mRNA โดยอาศัยลำดับเบสที่จำเพาะบนสาย mRNA ดังนั้นจึง
ไม่มีความจำเป็นที่จะต้องเติม m7G ทางปลายด้าน 5' อีกประการหนึ่งคือ การสังเคราะห์
mRNA และการสังเคราะห์โปรตีนเกิดในบริเวณเดียวกันหรือใกล้เคียงกันอีกทั้งยังเกิดขึ้นในเวลา
ไล่เลี่ยกันอีกด้วย ดังนั้นจึงไม่มีความจำเป็นที่จะต้องเติม poly-A tail ที่ปลายด้าน 3' เพื่อ
ป้องการถูกทำลายจากเอ็นไซม์ชนิดอื่น และเหตุผลสุดท้ายคือ pre-mRNA ของ โปรแคริโอต
ไม่มีอินทรอนจึงไม่เกิดการตัดแต่งในขั้นตอนที่ 3
|
: คุณคิดว่าความยาวของ poly-A tail ใน mRNA ต่างชนิดกันจะยาวเท่ากัน หรือไม่และถ้ายาวไม่เท่ากัน คุณคิดว่าน่าจะเป็นเพราะเหตุใด |
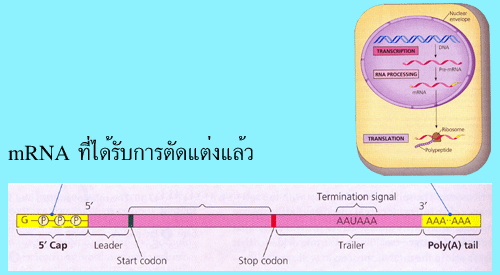
รูปที่ 3.11 mRNA ที่ได้รับการตัดแต่งแล้ว
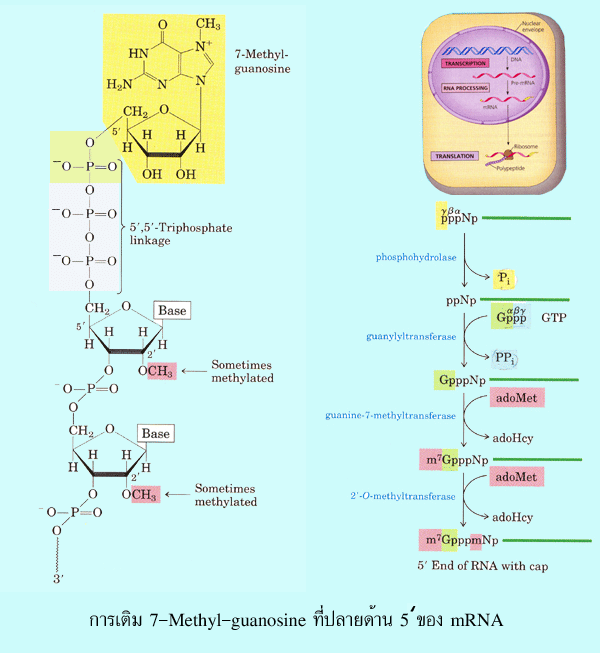
รูปที่ 3.12 การเติม 7-Methyl-guanosine ที่ปลายด้าน 5' ของ mRNA
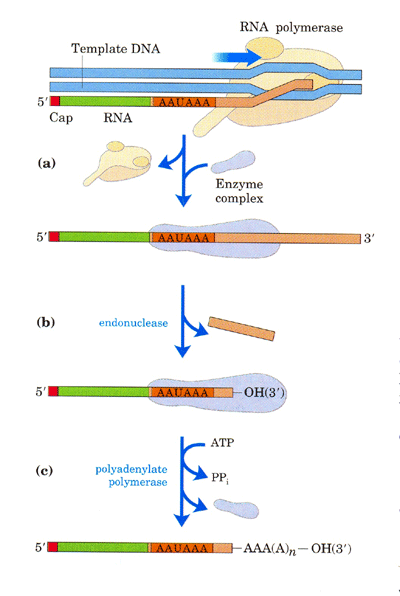
รูปที่ 3.13 การเติม poly-A tail ที่ปลายด้าน 3' ของ mRNA
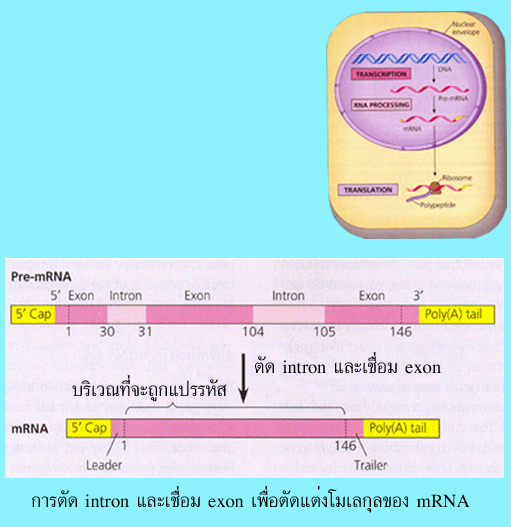
รูปที่ 3.14 การตัด intron และเชื่อม exon เพื่อตัดแต่งโมเลกุลของ mRNA

